Ciencia, Ambiente y Clima, Vol. 7, No. 2, julio-diciembre, 2024 ISSN (impreso): 2636-2317 • ISSN (en línea): 2636-2333 • Sitio web: https://revistas.intec.edu.do/
OCURRENCE OF IPS CALLIGRAPHUS (SCOLYTINAE: CURCULIONIDAE) IN REMNANT PINUS FOREST STANDS WITH VARYING FLORISTIC COMPOSITIONS AND DIFFERENT MANAGEMENT ON THE NORTHWESTERN SLOPE OF THE CENTRAL CORDILLERA, DOMINICAN REPUBLIC
Ocurrencia de Ips calligraphus (Scolytinae: Curculionidae) en rodales remanentes de bosques de Pinus con composiciones florísticas variables y diferente manejo en la ladera noroeste de la Cordillera Central, República Dominicana
ELÍ MISAEL BOBADILLA-PEÑALÓ1 Y ESCLAUDYS PÉREZ GONZÁLEZ2
Received: 21 de octubre 2024 • Accepted: 9 de enero 2025

How to cite: Bobadilla-Peñaló, E. M. & Pérez, E. (2024). Ocurrence of Ips calligraphus (Scolytinae: Curculionidae) in remnant Pinus forest stands with varying floristic compositions and different management on the northwestern slope of the Central Cordillera, Dominican Republic. Ciencia, Ambiente y Clima, 7(2), 49-73. https://doi.org/10.22206/cac.2024.v7i2.3346
Abstract
This study investigated the occurrence and population dynamics of Ips calligraphus within remnant Pinus forest stands with varying floristic compositions on the northwestern slope of the Cordillera Central, Dominican Republic. The study aimed to assess the influence of floristic diversity and forest management on the abundance of I. calligraphus in these fragmented ecosystems. Sampling was conducted across multiple forest stands with varying management and floristic compositions. A total of 18 plots of 20 x 20 meters were established, and I. calligraphus abundance was monitored using pheromone-baited traps over a five-week period. Tree dimensions, species richness, and floristic diversity were measured in each plot. Data analysis included descriptive statistics, diversity indices, and multivariate techniques such as Principal Component Analysis (PCA) and hierarchical clustering. The results indicated that I. calligraphus populations were more abundant in forest stands with higher floristic diversity, particularly in unmanaged sites. Significant variability in beetle populations was observed between managed and unmanaged forests, with unmanaged sites showing greater beetle pressure. PCA and clustering analyses revealed distinct capture profiles, suggesting that site-specific factors such as microclimate, forest management, and tree species composition influenced I. calligraphus dynamics. The study highlighted the complexity of managing I. calligraphus populations in fragmented pine forests. Floristic diversity, while beneficial for overall ecosystem health, did not appear to mitigate beetle infestations. The findings suggested the need for adaptive, site-specific pest management strategies that considered both ecological and management variables.
Keywords: natural forest stands, stablished forest stands, managed, unmanaged, diversity profile.
Resumen
Este estudio investigó la ocurrencia y dinámica poblacional de Ips calligraphus en remanentes de Pinus con diferentes composiciones florísticas en la vertiente noroeste de la Cordillera Central, República Dominicana. El objetivo del estudio fue evaluar la influencia de la diversidad florística y el manejo forestal en la abundancia de I. calligraphus en estos ecosistemas fragmentados. Se realizaron muestreos en varios rodales forestales con diferentes prácticas de manejo y composiciones florísticas. Se establecieron un total de 18 parcelas de 20 x 20 metros y se monitoreó la abundancia de I. calligraphus mediante trampas cebadas con feromonas durante un período de cinco semanas. En cada parcela se midieron las dimensiones de los árboles, la riqueza de especies y la diversidad florística. Los datos se analizaron mediante estadísticas descriptivas, índices de diversidad y técnicas multivariadas como el Análisis de Componentes Principales (PCA) y el análisis de agrupamiento jerárquico. Los resultados indicaron que las poblaciones de I. calligraphus eran más abundantes en los rodales con mayor diversidad florística, particularmente en los sitios no gestionados. Se observó una variabilidad significativa en las poblaciones de escarabajos entre los bosques gestionados y no gestionados, con una mayor presión de escarabajos en los sitios no gestionados. Los análisis de PCA y de agrupamiento revelaron perfiles de captura distintos, lo que sugiere que factores como el microclima, el manejo forestal y la composición de especies influyen en la dinámica de I. calligraphus. El estudio destacó la complejidad de gestionar las poblaciones de I. calligraphus en bosques de pinos fragmentados. Aunque la diversidad florística es beneficiosa para la salud general del ecosistema, no parece mitigar las infestaciones de escarabajos. Los hallazgos sugieren la necesidad de estrategias de manejo adaptativas y específicas para cada sitio, que consideren tanto los factores ecológicos como los de manejo.
Palabras clave: rodales forestales naturales, rodales forestales establecidos, gestionados, no gestionados, perfiles de diversidad.
Introduction
Ips calligraphus (Germar, 1823) commonly known as the six-spined engraver beetle, is a bark beetle belonging to the subfamily Scolytinae within the family Curculionidae (Coleoptera). Native to the Americas, I. calligraphus typically infests weakened, dying, or recently felled Pinus trees. Due to its ability to colonize and kill stressed or weakened pine trees, often in conjunction with associated blue-stain fungi (Pan et al., 2018; Goldazarena et al., 2012), their outbreaks can have significant ecological and economic consequences (Lundquist & Reich, 2014). The fragmentation of once-contiguous pine forests into smaller, isolated stands can have important implications for the dynamics of bark beetle populations (Barbosa & Marquet, 2002). Landscape heterogeneity has been shown to influence the dispersal patterns and population sizes of these insects, although the specific mechanisms are not yet fully understood (Lundquist & Reich, 2014).
In the Dominican Republic, the northwestern slope of the Central Cordillera presents a unique ecological niche characterized by remnant Pinus forest stands interspersed with varying floristic compositions. These forests have historically been impacted by outbreaks of I. calligraphus, particularly during periods of seasonal and prolonged drought (Jiménez et al., 2022). The study of I. calligraphus in these remnant forest stands is crucial for several reasons. First, the varying floristic compositions may influence the susceptibility of pine trees to infestations, affecting forest health and resilience. Second, understanding the occurrence patterns of this bark beetle can provide insights into the broader ecological dynamics and potential management strategies needed to preserve these forest ecosystems.
The forest stands on the northwestern slope of the Central Cordillera consist of both naturally established and planted Pinus forests. Naturally established forests typically emerge through ecological succession, where pioneer species gradually give way to more stable, mature communities. These forests often possess a higher degree of biodiversity and structural complexity, which can influence the distribution and impact of pest species like I. calligraphus. In contrast, planted Pinus forests are usually established through human intervention, such as reforestation projects or commercial timber operations, and may exhibit different ecological dynamics due to their often-uniform age structure and species composition.
The level of forest management also plays a critical role in the occurrence and impact of Ips (Hilszczański et al., 2007). Managed forests, where activities such as thinning, controlled burns, and pest monitoring are regularly conducted, tend to have better overall health and resilience against pest outbreaks (Marini et al., 2021). These practices can help reduce the population density of bark beetles and mitigate the severity of infestations. On the other hand, unmanaged forests, which are left to natural processes without human intervention, may be more vulnerable to pest outbreaks due to factors like increased tree density, accumulation of dead wood, and lack of active pest control measures (Kneeshaw et al., 2021).
The Central Cordillera’s northwestern slope, with its diverse topography and microclimates, creates a mosaic of habitats that support different plant species assemblages. This diversity may affect the population dynamics of I. calligraphus, either by providing refuge areas that reduce beetle pressure or by offering alternative hosts that sustain beetle populations. Based on this idea, this study aimed to investigate the occurrence of I. calligraphus in remnant Pinus forest stands with varying floristic compositions. By examining the relationship between beetle infestation rates and the diversity of plant species, as well as considering the establishment type (natural or planted) and management level (managed or unmanaged), we identified key factors that influence the persistence and spread of I. calligraphus.
Materials and Methods
Study Area
The study was conducted in the municipality of San José de las Matas (SAJOMA), located in the Santiago province of the Dominican Republic (Figure 1). This region is situated in the northwestern slope of Cordillera Central, within the coordinates of approximately 19.1833 N latitude and 70.9667 W longitude. San José de las Matas encompasses an area characterized by its diverse topography, which includes mountainous regions, rolling hills, and valley floors.
Figure 1
Study area and sampling sites
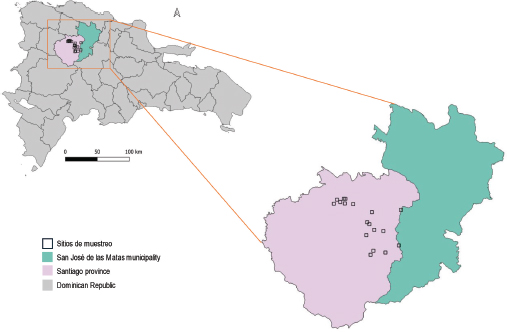
The elevation within the study area ranges from approximately 400 meters to over 900 meters above sea level (Table 1). The terrain is predominantly rugged, with steep slopes and deep valleys, contributing to varied microhabitats and ecological niches. SAJOMA´s climate is classified as tropical monsoon (Am) under the Köppen climate classification (Peel et al., 2007), with distinct wet and dry seasons. The average annual temperature is around 22°C, with temperatures typically ranging from 18°C to 28°C. The region experiences significant rainfall, with an average annual precipitation of approximately 1,800 mm. The wet season extends from May to November, characterized by heavy and frequent rains, while the dry season lasts from December to April.
Table 1
Study sites in Pinus spp remnants of northwestern slopes of Cordillera Central, Dominican Republic
Locality |
Pinus Specie |
Forest Remnant Stablisment |
Forest Management |
Coordenada |
Altitud (msnm) |
Bajamillo |
P. caribaea var. caribaea |
Planted |
Not managed |
19.27710, -70.95451 |
670 |
Guama |
P. caribaea var. caribaea |
Planted |
Managed |
19.35420, -70.84855 |
477 |
Las Carreras |
P. caribaea var. caribaea |
Planted |
Not managed |
19.23433, -70.89328 |
775 |
Montones Abajo |
P. caribaea var. caribaea |
Planted |
Not managed |
19.28232, -70.94997 |
575 |
Celestina |
P. caribaea var. hondurensis |
Planted |
Managed |
19.37024, -71.01443 |
525 |
La Mansión |
P. caribaea var. hondurensis |
Planted |
Managed |
19.34693, -70.93439 |
548 |
Corocito |
P. caribaea var. hondurensis |
Planted |
Managed |
19.23881, -70.92865 |
792 |
Celestina |
P. occidentalis |
Natural |
Managed |
19.36872, -70.98976 |
524 |
Mesquino |
P. occidentalis |
Natural |
Not managed |
19.29408, -70.89979 |
624 |
Rincón de Piedra |
P. occidentalis |
Planted |
Not managed |
19.22752, -70.93681 |
818 |
Damajagua |
P. occidentalis |
Natural |
Managed |
19.25467, -70.85371 |
741 |
The vegetation in San José de las Matas is diverse, reflecting the variation in altitude, climate, and soil types. The area hosts a range of habitats including tropical montane forests, pine forests, both natural and planted, and cloud forests. The lower elevations are dominated by tropical broadleaf forests and natural forest remnants of Pinus occidentalis and plantations of P. caribaea with different levels of management. The primary land use in San José de las Matas is agriculture and silviculture, with significant areas dedicated to the cultivation of coffee, citrics, and various fruits and vegetables. Livestock farming is also common. In recent years, eco-tourism has gained importance as a sustainable economic activity, promoting conservation and environmental awareness. However, the region faces challenges from deforestation, habitat fragmentation, and agricultural expansion, which pose threats to its biodiversity and ecological integrity.
Forest structure and associated flora
Eighteen 20 x 20 m (400 m²) plots were established to evaluate the flora and associated vegetation. Within each of these plots, two 5 x 5 m (25 m²) subplots and three 1 x 1 m (1 m²) subplots were placed to assess the shrub and herbaceous strata, respectively. All plots were square-shaped. In each plot, all plant species and their respective abundances were recorded. The structure of the tree and shrub strata was evaluated by considering all individuals with a diameter at breast height (DBH) ≥ 5 cm. The height of each individual was measured using a Suunto PM-5 forest clinometer. The measurement distance varied, depending on the distance from which the base and crown of the individual or a reference point could be clearly seen (Zobrist, 2008). Additionally, the DBH of each individual was measured at 1.30 m above the base using a Haglöf Mantax Blue caliper.
Sampling of bark beetles
Within each forest stand, artisanal traps baited with P361-Lure Ips calligraphus 1 Lure (Chemtica International S.A.) pheromone were established across the localities and forest remnant stands. The traps were placed at the exact center of each 400 m² square plot, mounted on stakes extending one meter above the ground. The number of individuals captured was quantified weekly, over a five-week period.
Data analysis
To determine the structure of forest remnants, descriptive statistical analysis of Diameter at Breast Height (DBH) and total height was conducted using the InfoStat statistical package (Di Rienzo et al., 2018). The effective number of species for Hill orders q = 0, 1, and 2 was calculated to assess species diversity (Chao et al., 2020). For these orders, sampling coverage and sample completeness were quantified. Rarefaction and extrapolation curves were used to predict the diversity expected with increased sampling, and most remnants showed reliable predictions. To compute the standard error and confidence intervals for the estimators, 100 bootstrap replications at a 95% confidence level were performed. Shannon and Simpson diversity indices were calculated to illustrate differences in species evenness and distribution further. Evenness estimates (Pielou’s J’, q = 1, q = 2) were used to evaluate biodiversity balance within the remnants, providing insights into the relative abundance and distribution of species and highlighting areas with well-balanced biodiversity versus those dominated by a few species. All richness and diversity analyses were performed using iNext4Steps Online (Chao & Hu, 2023). Principal Component Analysis (PCA) was employed to visualize and differentiate capture profiles across the localities, identifying distinct clusters of sites with similar environmental conditions or management practices. Hierarchical clustering analysis was performed for grouping sites with similar I. calligraphus captures.
Results
Forest remnants’ structure
The results on the Diameter at Breast Height (DBH) and Total Height of forest remnants of Pinus caribaea var. caribaea, Pinus caribaea var. hondurensis, and Pinus occidentalis (Table 2) highlight the complexity and diversity of Pinus forest remnants on the northwestern slopes of the Cordillera Central, particularly in a context where the ages of these natural and planted forest plots are not precisely known.
Table 2
Diameter at Breast Height (cm) and Total Height (m) of forest remnants of P. caribaea var. caribaea (Pcc), P. caribaea var. hondurensis (Pch), and P. occidentalis (Po)
Forest Remnant |
n |
Total Height (m) |
Diameter at Breast Height (cm) |
||||
Mean ± SD |
Min |
Max |
Mean ± SD |
Min |
Max |
||
Bajamillo (Pcc) |
10 |
14.65 ± 1.29 |
13 |
17 |
15.1 ± 3.86 |
9.24 |
19.75 |
Guama (Pcc) |
23 |
15.5 ± 1.86 |
12 |
19 |
19.41 ± 3.46 |
14.33 |
24.84 |
Las Carreras (Pcc) |
19 |
20.97 ± 1.53 |
16 |
23 |
30.84 ± 3.65 |
23.57 |
38.54 |
Montones Abajo (Pcc) |
40 |
16.34 ± 3.13 |
8.5 |
21 |
19.22 ± 5.13 |
11.78 |
28.34 |
Celestina (Pch) |
74 |
13.28 ± 4.9 |
3 |
22 |
19.66 ± 8.08 |
5.14 |
39.81 |
Corocito (Pch) |
10 |
16.6 ± 0.97 |
15 |
18 |
27.74 ± 2.08 |
22.93 |
30.25 |
La Mansión (Pch) |
18 |
17.28 ± 1.73 |
13 |
20 |
25.18 ± 3.83 |
17.83 |
30.57 |
Celestina (Po) |
94 |
13.7 ± 5.25 |
2.5 |
21.5 |
17.27 ± 8.49 |
5.46 |
38.22 |
Damajagua (Po) |
10 |
11.55 ± 1.07 |
10 |
13 |
22.2 ± 4.26 |
17.2 |
32.48 |
Mesquino (Po) |
13 |
17.62 ± 2.58 |
12 |
21 |
18.52 ± 5.84 |
8.28 |
27.71 |
Rincón de Piedra (Po) |
14 |
16.43 ± 5.75 |
6 |
22 |
20.81 ± 9.88 |
5.41 |
33.12 |
P. caribaea var. caribaea (Pcc), P. caribaea var. hondurensis (Pch), and P. occidentalis (Po).
P. caribaea var. caribaea exhibited considerable variation in both total height and DBH across different forest remnants, influenced by their management and establishment status. For instance, Las Carreras, a planted and unmanaged remnant, recorded the highest mean total height and the largest mean DBH (Table 2). This suggests that even without active management, Las Carreras, a traditional area for cultivating forest species, provides favorable growth conditions for P. caribaea var. caribaea, possibly due to microclimatic conditions or lower levels of disturbance. Bajamillo, another planted but unmanaged site, had a lower mean total height and DBH (Table 2). The substantial standard deviation in DBH at Montones Abajo (SD = 5.13), also a planted and unmanaged site, points to significant heterogeneity in tree sizes, reflecting a mixed-age forest structure within the remnant due to constant replanting as a forest management strategy in response to tree losses caused by I. calligraphus. In managed remnants like Guama the managed status could contribute to more consistent growth conditions, resulting in less variability compared to unmanaged sites (Table 2).
P. caribaea var. hondurensis showed notable variability in both height and DBH in its planted and managed remnants. The Celestina site had a mean total height with a substantial standard deviation, indicating a wide range of tree heights from (Table 2). This high variability could be attributed to different growth rates among trees, possibly due to variations in light availability, soil nutrients, past disturbances caused by forest fires, and infestations of I. calligraphus. Similarly, the mean DBH at Celestina suggests a wide range of tree diameters, further supporting the notion of a heterogeneous forest structure. In contrast, Corocito, another planted and managed site, had more uniform tree sizes, with a relatively low standard deviation (Table 2), indicating more consistent growth conditions or a more uniform age class of trees.
P. occidentalis showed distinct structural characteristics across its natural and planted remnants. The Damajagua site, a natural and managed remnant, had the lowest mean total height and a relatively low standard deviation (Table 2), suggesting a population of shorter and possibly younger trees. On the other hand, the Celestina site, a natural and managed remnant, showed a high degree of variability in both total height and DBH (Table 2), indicating a complex forest structure with a wide range of tree sizes. This variability could be due to diverse microhabitats, historical land use, or differentiated regeneration dynamics, particularly in the various historical scenarios of I. calligraphus attacks that this locality has suffered. The Rincón de Piedra site, a planted and unmanaged remnant, had substantial variability for both, height and DBH (Table 2), reflecting a heterogeneous environment possibly influenced by varied topography, soil conditions, or the lack of management practices caused by the abandonment of these forest plots.
Species richness and diversity of flora associated with forest remnants
Table 3 provides a comprehensive overview of biodiversity and sampling effectiveness across the forest remnants, offering insights into observed species richness and sample coverage. Additionally, it includes metrics for sample completeness (q = 0, q = 1, q = 2), providing a more nuanced understanding of sampling adequacy and biodiversity representation.
Table 3
Sampling coverage and sample completeness for the forest remnants of P. caribaea var. caribaea, P. caribaea var. hondurensis, and P. occidentalis
Forest Remnant |
n |
Sampling coverage |
Sample Completeness |
|||||||||
S.obs |
SC(n) |
SC(2n) |
f1 |
f2 |
f3 |
f4 |
f5 |
q = 0 |
q = 1 |
q = 2 |
||
Bajamillo (Pcc) |
203 |
5 |
0.98 |
0.99 |
4 |
0 |
0 |
0 |
0 |
0.46 |
0.98 |
1.00 |
Montones Abajo (Pcc) |
195 |
22 |
0.99 |
1.00 |
2 |
3 |
1 |
3 |
2 |
0.91 |
0.98 |
1.00 |
Las Carreras (Pcc) |
11 |
2 |
1.00 |
1.00 |
0 |
0 |
1 |
0 |
0 |
0.93 |
0.98 |
1.00 |
Guama (Pcc) |
66 |
15 |
0.96 |
1.00 |
3 |
5 |
4 |
1 |
1 |
1.00 |
1.00 |
1.00 |
Celestina (Pch) |
250 |
21 |
0.98 |
1.00 |
4 |
4 |
1 |
2 |
1 |
0.80 |
0.96 |
1.00 |
Corocito (Pch) |
14 |
3 |
1.00 |
1.00 |
1 |
0 |
0 |
1 |
0 |
0.94 |
0.96 |
0.99 |
La Mansión (Pch) |
81 |
14 |
0.95 |
0.97 |
4 |
1 |
2 |
1 |
1 |
0.64 |
0.95 |
1.00 |
Celestina (Po) |
204 |
26 |
0.98 |
1.00 |
5 |
6 |
2 |
4 |
2 |
1.00 |
1.00 |
1.00 |
Damajagua (Po) |
48 |
4 |
0.96 |
0.99 |
2 |
0 |
0 |
0 |
0 |
0.55 |
0.78 |
0.97 |
Mesquino (Po) |
13 |
5 |
0.78 |
0.89 |
3 |
1 |
0 |
0 |
0 |
0.97 |
0.99 |
1.00 |
Rincón de Piedra (Po) |
92 |
9 |
1.00 |
1.00 |
1 |
0 |
2 |
1 |
0 |
1.00 |
1.00 |
1.00 |
n = number of observed individuals in the reference sample (sample size). S.obs = number of observed species in the reference sample. SC(n) = sample coverage estimates of the reference sample. SC(2n) = sample coverage estimates for twice the reference sample size. f1-f5 = the first five species abundance frequency counts in the reference sample.
The data highlights significant variability in the number of observed individuals, ranging from as few as 11 in Las Carreras (Pcc) to as many as 250 in Celestina (Pch). Despite these differences, most forest remnants exhibit high sample coverage estimates close to 1, indicating that the samples are nearly comprehensive in capturing the species present. For example, Bajamillo (Pcc) with 203 individuals has an SC(n) of 0.98 and SC(2n) of 0.99, demonstrating that a substantial portion of the species diversity has been captured even with the current sample size. Similarly, Montones Abajo (Pcc) and Corocito (Pch) exhibit high sample coverage, suggesting effective sampling strategies.
The first five species abundance frequency counts (f1 to f5) reveal the distribution of species abundances within each remnant. For instance, Celestina (Po) shows a high frequency of species in the first five abundance counts, indicating a relatively even species distribution. In contrast, Damajagua (Po) has a lower number of species and sample completeness (q = 0.55), highlighting a less diverse ecosystem. The sample completeness metrics (q values) provide further insight, with most remnants showing high completeness (q = 1 and q = 2), affirming the thoroughness of the sampling efforts.
The rarefaction curves (solid lines) provide insights into the observed diversity up to the current sample size, while the extrapolation curves (dashed lines) predict the diversity that would be observed with increased sampling (Figure 2). Most forest remnants show a smooth transition from rarefaction to extrapolation, indicating reliable predictions of diversity with further sampling. Notably, remnants such as Montones Abajo (Pcc) and Celestina (Po) show little change in diversity estimates beyond their current sample sizes, suggesting that most of their diversity has been captured. Conversely, remnants like La Mansión (Pch) and Mesquino (Po) exhibit more pronounced extrapolation curves, indicating the potential for discovering additional species with further sampling.
Figure 2
Sample size-based rarefaction and extrapolation sampling curve for the flora associated with the forest remnants
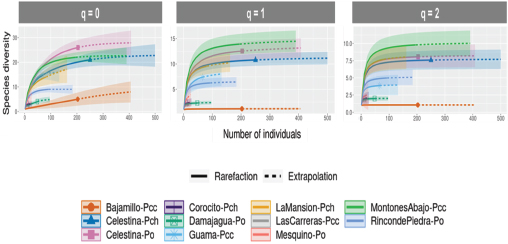
In terms of species richness, Montones Abajo (Pcc) and Celestina (Po) exhibit the highest diversity, leveling off as the sample size approaches 500 individuals, suggesting high initial diversity that is quickly captured with increased sampling. In contrast, remnants like Bajamillo (Pcc) and Las Carreras (Pcc) show much lower species richness, indicating less diverse ecosystems.
Shannon diversity curves reveal that Montones Abajo (Pcc) and Guama (Pcc) have high diversity, suggesting a rich and evenly distributed community. Conversely, remnants like Damajagua (Po) and Las Carreras (Pcc) exhibit lower Shannon diversity, reflecting fewer even species distributions. Simpson diversity curves indicate that Montones Abajo (Pcc) and Celestina (Po) have high evenness and low species dominance, while remnants like Mesquino (Po) and Damajagua (Po) have lower diversity, indicating a more uneven species distribution. The combination of rarefaction and extrapolation in these curves shows reliable predictions of diversity with further sampling, highlighting the varying biodiversity and sampling adequacy across different forest remnants.
Table 4 offers a detailed examination of the taxonomic diversity within forest remnants, The observed values (TD_obs) are compared against the estimated asymptotic values (TD_asy), with bootstrap standard errors and confidence limits providing further context on the reliability and comprehensiveness of the sampling efforts.
Table 4
Observed and estimated taxonomic diversity across forest remnants
Forest Remnant |
qTD |
TD_obs |
TD_asy |
s.e. |
qTD.LCL |
qTD.UCL |
Bajamillo (Pcc) |
q = 0: Species richness |
5 |
10.97 |
3.15 |
4.8 |
17.14 |
q = 1: Shannon diversity |
1.13 |
1.16 |
0.07 |
1.03 |
1.29 |
|
q = 2: Simpson diversity |
1.04 |
1.04 |
0.02 |
1 |
1.08 |
|
Guama (Pcc) |
q = 0: Species richness |
15 |
15.89 |
3.58 |
8.87 |
22.91 |
q = 1: Shannon diversity |
7.38 |
8.37 |
1.28 |
5.86 |
10.87 |
|
q = 2: Simpson diversity |
3.88 |
4.05 |
0.86 |
2.38 |
5.73 |
|
Las Carreras (Pcc) |
q = 0: Species richness |
2 |
2 |
0.2 |
1.61 |
2.39 |
q = 1: Shannon diversity |
1.8 |
1.89 |
0.26 |
1.38 |
2.39 |
|
q = 2: Simpson diversity |
1.66 |
1.77 |
0.33 |
1.12 |
2.43 |
|
Montones Abajo (Pcc) |
q = 0: Species richness |
22 |
22.66 |
2.93 |
16.93 |
28.4 |
q = 1: Shannon diversity |
13.91 |
14.76 |
0.94 |
12.91 |
16.61 |
|
q = 2: Simpson diversity |
9.79 |
10.26 |
1.01 |
8.28 |
12.24 |
|
Celestina (Pch) |
q = 0: Species richness |
21 |
22.99 |
6.01 |
11.22 |
34.76 |
q = 1: Shannon diversity |
10.79 |
11.32 |
0.73 |
9.89 |
12.76 |
|
q = 2: Simpson diversity |
7.59 |
7.8 |
0.63 |
6.57 |
9.02 |
|
Corocito (Pch) |
q = 0: Species richness |
3 |
3 |
0.52 |
1.98 |
4.02 |
q = 1: Shannon diversity |
2.29 |
2.48 |
0.4 |
1.7 |
3.26 |
|
q = 2: Simpson diversity |
2 |
2.17 |
0.46 |
1.26 |
3.08 |
|
La Mansión (Pch) |
q = 0: Species richness |
14 |
21.9 |
8.62 |
5.01 |
38.79 |
q = 1: Shannon diversity |
9.53 |
10.87 |
1.32 |
8.29 |
13.45 |
|
q = 2: Simpson diversity |
7.69 |
8.39 |
0.95 |
6.53 |
10.26 |
|
Celestina (Po) |
q = 0: Species richness |
26 |
28.07 |
6.74 |
14.87 |
41.28 |
q = 1: Shannon diversity |
12.54 |
13.49 |
1.11 |
11.32 |
15.66 |
|
q = 2: Simpson diversity |
8.06 |
8.35 |
0.79 |
6.8 |
9.9 |
|
Damajagua (Po) |
q = 0: Species richness |
4 |
4.98 |
0.57 |
3.87 |
6.09 |
q = 1: Shannon diversity |
2.27 |
2.39 |
0.21 |
1.98 |
2.79 |
|
q = 2: Simpson diversity |
1.99 |
2.03 |
0.19 |
1.67 |
2.4 |
|
Mesquino (Po) |
q = 0: Species richness |
5 |
9.15 |
4.13 |
1.07 |
17.24 |
q = 1: Shannon diversity |
3.25 |
4.6 |
1.86 |
0.96 |
8.24 |
|
q = 2: Simpson diversity |
2.38 |
2.69 |
1.12 |
0.5 |
4.88 |
|
Rincón de Piedra (Po) |
q = 0: Species richness |
9 |
9 |
0.93 |
7.18 |
10.82 |
q = 1: Shannon diversity |
6.24 |
6.53 |
0.49 |
5.57 |
7.49 |
|
q = 2: Simpson diversity |
4.97 |
5.2 |
0.51 |
4.21 |
6.19 |
qTD = ‘Species richness’ represents the diversity of order q=0; ‘Shannon diversity’ represents the diversity of order q=1, ‘Simpson diversity’ represents the diversity of order q=2. TD_obs = the empirical diversity value of order q. TD_asy = the estimated asymptotic diversity value of order q. s.e. = the bootstrap standard error of the estimated asymptotic diversity of order q. qTD.LCL, qTD.UCL = the bootstrap lower and upper confidence limits for the estimated asymptotic diversity of order q at the specified level in the setting (with a default value of 0.95). Pielou J’ = a widely used evenness measure and it is expressed as J’ = H/log(S) where H denotes Shannon entropy.
The analysis reveals considerable variability in species richness across the remnants. For instance, Bajamillo (Pcc) shows an observed species richness of 5, but the estimated asymptotic richness suggests there could be up to 10.97 species. Similarly, Celestina (Po) has a high observed richness of 26, with an asymptotic estimate of 28.07, suggesting that the sampling was comprehensive, though there may still be more species to uncover. The confidence intervals provide a range within which the true species richness likely falls, highlighting the potential for additional species discovery in these ecosystems.
The diversity measures further illustrate the differences in species evenness and distribution. Montones Abajo (Pcc) displays a high Shannon diversity of 13.91 and an estimated value of 14.76, indicating a diverse and evenly distributed community. In contrast, remnants like Mesquino (Po) exhibit lower diversity and higher variability, with a Shannon diversity of 3.25 and an asymptotic estimate of 4.6, reflecting possible under-sampling and uneven species distribution. Simpson diversity values similarly underscore the varying degrees of species dominance, with Guama (Pcc) and Rincón de Piedra (Po) showing higher evenness compared to remnants like Damajagua (Po). Overall, the table underscores the importance of comprehensive sampling to accurately capture the biodiversity within these forest remnants. The observed and estimated diversity metrics reveal significant insights into species richness and evenness, guiding conservation efforts and highlighting areas where further sampling may be needed. This analysis not only enhances our understanding of the ecological dynamics within these remnants but also informs strategies for biodiversity preservation and management.
The evenness estimates reveal critical insights into the biodiversity and ecological balance of the forest remnants (Table 5). Bajamillo (Pcc) stands out for its low evenness across all measures, indicating that it is dominated by a few species with little representation of others. This suggests that Bajamillo might be an ecosystem under stress or with specific dominant species outcompeting others. On the other hand, remnants like Montones Abajo (Pcc), La Mansión (Pch), and Rincón de Piedra (Po) exhibit high evenness, indicating a healthier and more balanced ecosystem where species are more equally represented. This balance is crucial for the stability and resilience of these ecosystems, as it suggests a lower likelihood of any single species dominating and potentially disrupting ecological processes.
Table 5
Evenness estimates of orders q = 1 and 2 at the standardized coverage value of Cmax based on the normalized slope of a diversity profile
Cmax = 0.892 |
Pielou’J |
q = 1 |
q = 2 |
Bajamillo (Pcc) |
0.14 |
0.11 |
0.04 |
Guama (Pcc) |
0.74 |
0.47 |
0.22 |
Montones Abajo (Pcc) |
0.87 |
0.68 |
0.49 |
Las Carreras (Pcc) |
0.82 |
0.78 |
0.62 |
Celestina (Pch) |
0.86 |
0.68 |
0.50 |
Corocito (Pch) |
0.82 |
0.76 |
0.60 |
La Mansión (Pch) |
0.89 |
0.75 |
0.60 |
Celestina (Po) |
0.82 |
0.56 |
0.37 |
Damajagua (Po) |
0.84 |
0.79 |
0.63 |
Mesquino (Po) |
0.70 |
0.48 |
0.25 |
Rincón de Piedra (Po) |
0.87 |
0.75 |
0.59 |
The measures for q=1 and q=2 further differentiate between remnants by considering species abundance and dominance, respectively. Higher values for these measures indicate ecosystems where biodiversity is maintained without dominance by a few species, which is a positive indicator of ecological health. In conclusion, these evenness estimates provide valuable information for conservation efforts, highlighting areas where biodiversity is well-balanced and areas that might require more targeted management to improve species distribution and ecosystem stability.
I. Calligraphus abundance across remnants
I. calligraphus weekly captures (Table 6) highlights considerable variation in species counts across different forest remnants over the five-week period. Montones Abajo (Pcc) and Rincón de Piedra (Po) stand out for their high total and median counts, indicating robust species presence. However, the high median absolute deviation (MAD) values for these remnants suggest significant weekly fluctuations, which could be due to various ecological factors or sampling inconsistencies. In contrast, La Celestina (Po), despite having lower total counts, shows minimal weekly variation, reflected in its low MAD value. This stability could indicate a more balanced ecosystem.
Table 6
Weekly captures of I. calligraphuss in forest remnants
Locality (Stand’s Specie) |
Week 1 |
Week 2 |
Week 3 |
Week 4 |
Week 5 |
Total |
Median |
MAD |
Bajamillo (Pcc) |
231 |
182 |
108 |
148 |
43 |
712 |
148 |
40 |
Guama (Pcc) |
14 |
7 |
5 |
14 |
17 |
57 |
14 |
3 |
Las Carreras (Pcc) |
126 |
116 |
92 |
114 |
83 |
531 |
116 |
22 |
Montones Abajo (Pcc) |
213 |
238 |
448 |
448 |
318 |
1665 |
318 |
105 |
La Celestina (Pch) |
42 |
86 |
57 |
31 |
63 |
279 |
57 |
14 |
La Mansión (Pch) |
26 |
56 |
42 |
11 |
92 |
227 |
42 |
21.5 |
Corocito (Pch) |
147 |
94 |
240 |
121 |
213 |
815 |
147 |
53 |
La Celestina (Po) |
43.5 |
40 |
48.5 |
51 |
42 |
225 |
43.5 |
4.5 |
Mesquino (Po) |
329 |
32 |
78 |
46 |
54 |
539 |
78 |
32 |
Rincón de Piedra (Po) |
411 |
178 |
162 |
185 |
35 |
971 |
178 |
64 |
Damajagua (Po) |
147 |
64 |
36 |
19 |
53 |
319 |
53 |
19 |
The Principal Component Analysis (PCA) scatter plot (Figure 3) provides a comprehensive view of the I. calligraphus capture profiles across the localities. The plot reveals distinct clusters of sites, indicating shared environmental conditions or management practices that influence I. calligraphus populations. For instance, Montones Abajo and Corocito are closely grouped in the upper right quadrant, suggesting that they exhibit similar capture profiles. Bajamillo (Pcc) and Rincón de Piedra (Pcc), while in the same general area, show slight differences, indicating some unique characteristics compared to Montones Abajo and Corocito (Pch). In contrast, La Mansión (Pch), Celestina (Pch, Po), and Guama (Pcc) form a tight cluster in the lower left quadrant, reflecting very similar capture profiles, whereas Damajagua (Po), though in the same quadrant, shows some variance.
Figure 3
Principal Component Analysis scatter plot for the forest remnants
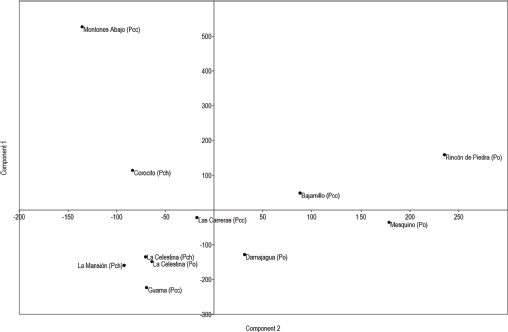
The PCA also identifies outliers such as Mesquino (Po), positioned distinctly in the lower right quadrant, and Las Carreras (Pcc), which is centrally located but slightly apart from the other clusters. These outliers suggest unique capture profiles that may be influenced by different environmental conditions or specific management practices. Component 1, which captures the largest variance, highlights the significant differences in capture profiles between sites like Montones Abajo (Pcc) and Rincón de Piedra (Po) compared to La Celestina (Pch, Po) and Guama (Pcc). Component 2 further distinguishes Mesquino (Po)’s unique characteristics, underscoring the need for specialized management practices.
Overall, the PCA scatter plot underscores the importance of understanding site-specific factors in managing I. calligraphus populations. Locations within the same cluster may benefit from similar management strategies or environmental conditions, while outliers such as Mesquino (Po) require tailored approaches. Further investigation into the specific conditions driving these differences can enhance the effectiveness of pest management, ensuring that strategies are appropriately targeted to the unique characteristics of each site.
The hierarchical clustering analysis of I. calligraphus capture profiles across the forest remnants (Figure 4) reveals distinct groupings that reflect underlying ecological similarities and differences. The dendrogram shows two major clusters, indicating that certain locations share similar environmental conditions or management practices that result in comparable I. calligraphus populations. The first major cluster includes Las Carreras (Pcc), Corocito (Pch), Mesquino (Po), Rincón de Piedra (Po), Bajamillo (Pcc), and Montones Abajo (Pcc). Within this cluster, Montones Abajo (Pcc) and Rincón de Piedra (Po) are particularly similar, suggesting that these sites could potentially be managed with similar strategies for I. calligraphus control. Mesquino (Po), while part of this cluster, stands somewhat apart, indicating unique conditions that may require tailored management practices despite its overall similarity to the group.
Figure 4
Hierarchical clustering analysis of I. calligraphus capture profiles across the forest remnants
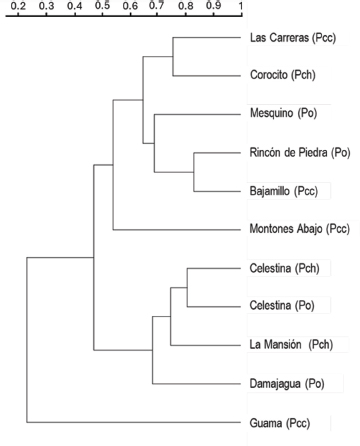
The second major cluster comprises Damajagua (Po), Las Carreras (Pcc), La Celestina, La Mansión (Pch), and Guama (Pcc). Damajagua (Po) and Las Carreras (Pcc) form a closely related sub-cluster, suggesting very similar capture profiles and potentially interchangeable management strategies. La Celestina (Pch, Po), La Mansión (Pch), and Guama (Pcc), though part of the same larger cluster, display a more nuanced relationship, with La Celestina (Pch, Po) and La Mansión (Pch) being more closely related to each other than to Guama (Pcc). This hierarchical structure underscores the importance of understanding site-specific conditions that influence I. calligraphus populations, guiding targeted pest management interventions.
Discussion
The variability in mean height and DBH observed among forest remnants highlights the influence of forest management and establishment on tree growth and overall forest structure. Sites like Las Carreras, which are planted but unmanaged, exhibited significant variability in DBH and tree height. This suggests that a lack of consistent management practices, such as thinning or pest control, might lead to mixed-age structures, where replanting occurs as a response to pest-induced tree losses. This aligns with findings by Hilszczański et al. (2007)), who noted that unmanaged forests could be more susceptible to pest outbreaks due to higher tree density and accumulation of dead wood, providing ample habitat for beetle colonization.
On the other hand, managed sites like Guama and Corocito showed more uniform tree sizes, which can be attributed to regular management activities, including thinning, weeding or even controlled burns. Such practices help reduce competition, improve tree vigor, and ultimately lower the susceptibility of pines to beetle infestations (Marini et al., 2021). However, even within managed sites, I. calligraphus populations were still present, indicating that management reduces but does not entirely prevent infestations. This suggests that other ecological factors, such as floristic diversity and microhabitat conditions, also play a significant role in beetle dynamics (Kneeshaw et al., 2021).
One of the key findings of this study is the relationship between floristic diversity and I. calligraphus abundance. The results showed that forest stands with more diverse understory and midstory vegetation tended to support higher beetle populations. For instance, Montones Abajo and La Mansión, both with higher diversity indices, had robust I. calligraphus captures. This finding challenges the initial hypothesis that increased diversity would create a more complex habitat, thereby reducing beetle colonization. Instead, it appears that diverse forests may provide microhabitats or stress factors that could favor beetle presence (Pan et al., 2018).
The variability in beetle captures across sites suggests that floristic diversity may contribute to beetle persistence by creating a mosaic of conditions where some plants might stress the pines, making them more susceptible to infestation. Additionally, the presence of other plant species might serve as alternative hosts or support associated fungi that aid beetle survival, a phenomenon observed in other studies (Goldazarena et al., 2012). This implies that while floristic diversity can enhance ecosystem health, it does not necessarily deter pests like I. calligraphus, indicating a complex interplay between plant diversity and pest dynamics.
The analysis of forest remnants revealed that species-specific traits and site-specific conditions also affect I. calligraphus populations. For example, P. caribaea var. caribaea at Las Carreras, despite being unmanaged, showed substantial growth and high beetle presence, possibly due to favorable microclimatic conditions that mitigate the stress from beetle attacks. Conversely, managed sites like Corocito, which had consistent tree sizes, showed a less dramatic but still notable presence of beetles, suggesting that even well-managed forests are not immune to beetle infestation (Jiménez et al., 2022). Sites like Montones Abajo and Rincón de Piedra clustered together (Figure 4), indicating shared ecological conditions that favored high beetle populations. This clustering suggests that management strategies could be effective across these sites. However, outliers such as Mesquino showed unique capture profiles, which may be influenced by specific environmental conditions, such as soil type, microclimate, or past disturbances (Lundquist & Reich, 2014). These variations highlight the importance of localized management approaches that address site-specific conditions rather than applying broad, uniform strategies.
The study’s findings underscore the need for adaptive management strategies that are informed by detailed ecological assessments. Regular monitoring of I. calligraphus populations, combined with data on tree health, floristic composition, and environmental variables, can help in developing targeted pest control measures. For instance, in sites where high beetle abundance is linked to specific plant associations, removing or managing those understory plants could be explored as a control method. Moreover, consistent management practices like thinning, weeding or even controlled burns, as seen in Guama, appear to mitigate beetle outbreaks by reducing tree competition and maintaining forest vigor (Marini et al., 2021). However, the study also reveals that such practices must be continuous; sporadic management may lead to transient reductions in beetle populations, but not sustainable control. Additionally, understanding how floristic diversity influences beetle dynamics can help forest managers design plantings that are both ecologically rich and resilient against pest infestations, perhaps by selecting companion species that do not favor beetle persistence.
The study provides a comprehensive view of how I. calligraphus populations are influenced by various ecological and management factors within the remnant forests of the northwestern slopes of the Cordillera Central. While floristic diversity, management practices, and forest structure each play significant roles, their interactions create complex dynamics that can either mitigate or exacerbate beetle infestations. Effective pest management will therefore require adaptive strategies that consider these factors, focusing on enhancing forest health and resilience through a combination of regular monitoring, tailored management practices, and site-specific interventions.
These findings align with broader ecological research on bark beetles, emphasizing the importance of landscape heterogeneity, continuous management, and understanding specific plant-pest interactions in preventing widespread outbreaks. Future studies could explore more deeply the mechanisms by which plant diversity and microhabitat conditions contribute to beetle dynamics, offering further insights for sustainable forest management.
Funding
This research was made possible thanks to the financial support of the National Fund for Innovation in Scientific and Technological Development (Fondocyt), through project No. VCT-FONDOCYT-001, Call 2022, titled “Molecular characterization of fungal communities associated with the bark beetle Ips calligraphus, their interrelation in the death of Pinus occidentalis, P. caribaea, and P. caribaea var. hondurensis, and evaluation of biocontrol measures for the Ips calligraphus-fungus complex.”
Acknowledgments
Thanks, are also extended to Plan Sierra Inc. for their logistical support during the fieldwork, provided through their Research Unit, which facilitated the execution of this study.
References
Barbosa, O., & Marquet, P. (2002). Effects of forest fragmentation on the beetle assemblage at the relict forest of Fray Jorge, Chile. Oecologia, 132, 296-306. https://doi.org/10.1007/s00442-002-0951-3.
Chao, A. & Hu, K.-H. (2023). iNEXT.4steps Online: four-steps biodiversity analysis based on iNEXT. Code and user’s guide available from https://chao.shinyapps.io/iNEXT_4steps
Chao, A., Kubota, Y., Zelený, D., Chiu, C.-H., Li, C.-F., Kusumoto, B., Yasuhara, M., Thorn, S., Wei, C.-L., Costello, M. J. and Colwell, R. K. (2020). Quantifying sample completeness and comparing diversities among assemblages. Ecological Research, 35, 292-314.
Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, and, M., Robledo C.W. (2018). InfoStat versión 2018. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
Goldarazena, A., Romon Ochoa, P., & López, S. (2012). Bark beetles control in forests of northern Spain. In Insecticides - Pest Engineering (pp. 323-352). InTech. https://doi.org/10.5772/3016
Hilszczański, J., Gibb, H., & Bystrowski, C. (2007). Insect natural enemies of Ips typographus (L.) (Coleoptera, Scolytinae) in managed and unmanaged stands of mixed lowland forest in Poland. Journal of Pest Science, 80, 99-107. https://doi.org/10.1007/s10340-006-0160-7.
Jiménez G., A. A., Cordón Cabrera, P. R., & Agramonte, J. E. (2022). Fluctuación poblacional de Ips calligraphus en pinares de la vertiente norte de la cordillera Central, República Dominicana. Novitates Caribaea, (20), 137–154. https://doi.org/10.33800/nc.vi20.314
Kneeshaw, D., Sturtevant, B., De Grandpé, L., Doblas-Miranda, E., James, P., Tardif, D., & Burton, P. (2021). The Vision of Managing for Pest-Resistant Landscapes: Realistic or Utopic? Current Forestry Reports, 7, 97 - 113. https://doi.org/10.1007/s40725-021-00140-z.
Marini, L., Ayres, M., & Jactel, H. (2021). Impact of Stand and Landscape Management on Forest Pest Damage. Annual Review of Entomology. https://doi.org/10.1146/annurev-ento-062321-065511.
Lundquist, J. E., & Reich, R M. (2014). Landscape Dynamics of Mountain Pine Beetles. Oxford University Press, 60(3), 464-475. https://doi.org/10.5849/forsci.13-064
Pan, Y., Zhao, T., Krokene, P., Yu, Z., Qiao, M., Lü, J., Chen, P., & Ye, H. (2018). Bark Beetle-Associated Blue-Stain Fungi Increase Antioxidant Enzyme Activities and Monoterpene Concentrations in Pinus yunnanensis. Frontiers Media, 9. https://doi.org/10.3389/fpls.2018.01731
Peel M.C., Finlayson B.L. & McMahon T.A. (2007). Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci., 11, 1633–1644.
Zobrist, K. W. (2008). Virtual cruiser vest: measuring your forest stand. Recuperado 2 de julio de 2022, De http://www.ruraltech.org/virtual_cruiser/contacts/index.html
_______________________________
1 Universidad ISA. Ave. Pres. Antonio Guzmán Km. 5 1/2, La Herradura, Santiago. Apartado Postal 166. ORCID: 0000-0002-0725-9854. Correo-e: ebobadilla@isa.edu.do
2 Universidad ISA. Ave. Pres. Antonio Guzmán Km. 5 1/2, La Herradura, Santiago. Apartado Postal 166. ORCID: 0000-0003-0465-6261. Correo-e: eperez@isa.edu.do