Ciencia, Ambiente y Clima, Vol. 5, No. 1, enero-junio, 2022 • ISSN (impreso): 2636-2317 • ISSN (en línea): 2636-2333 • Sitio web: https://revistas.intec.edu.do/
CONTRIBUTION TO COMPETING SPECIES MODELING WITH DISTRIBUTED DELAY
Contribución a los Modelos de Especies Competidoras con Retardo Distribuido
* Universidad de Oriente. Cumaná, Venezuela. ORCID: https://orcid.org/0000-0002-2482-4253, Correo-e: mjcavani@gmail.com
Recibido: ● Aprobado:

Cómo citar: Cavani, M. (2022). Contribution to Competing Species Modeling with Distributed Delay. Ciencia, Ambiente y Clima, 5(1), 85–95. https://doi.org/10.22206/cac.2022.v5i1.pp85-95
1. Introduction
Hsu, Hubbell and Waltman have analyzed the behavior of a predator-prey system consisting of two predators' species x1 , x2 and a single prey specie, S (1978a, 1978b). The predator species compete purely exploitative, with no interference between rivals (no toxins are produced, for example). Both species have access to the prey and compete only by lowering the population of shared prey. For death rates it is assumed that the number dying is proportional to the number currently alive. In the absence of predation, the prey grows logistically, and the predator’s functional response obeys the Michaelis-Menten kinetic, which is also called the Holling type curve. Thus, it is assumed that if there are no significant time lags in the system, the following model may describe the situation:
Here: xi(t) is the number of the i-th predator at time t; S(t) is the number of the prey at time t; mi is the maximum growth rate of the i−th predator; Di is the death rate for the i-th predator; ai is the half-saturation constant for the i-th predator, which is the prey density at which the functional response of the predator is half maximal; γ and K are the intrinsic rate of increase and the carrying capacity for the prey population respectively. The model (1) has been extensively studied for several authors, nonetheless here a distributed delay is introduced and using the linear trick chain technique the analysis of the dynamics of the equilibria and the main properties of the model are studied. The delay system is converted by mean of linear trick chain technique in an “equivalent system” of five ordinary differential equations. The final purpose is to look for conditions where neither, one, or both species of predators survive.
2. Formulation of the model
As it was mentioned early, a modification of the predator-prey model (1) will be performed. The main consideration is that there are lags in transforming the consumed prey biomass into new biomass of the predator populations. In order to model this process of conversion of prey consumed into predators, a distributed delay is introduced in the system to describe the involved time lag. More precisely, assume in a more realistic fashion that the present level of the predators affects instantaneously the growth of the prey, but the growth of the predator is influenced by the amount of prey in the past. Thus, suppose that the predator grows up depending on the weight average time of the function of Michaelis-Menten of the prey S over the past per predator. Thus, the model takes the form of the following integro-differential system:
In the literature, the kernel
(2)
is called the weak kernel and is frequently used in biological modeling and clearly implies that the influence of the past is fading away exponentially and the number 1/αi might be interpreted as the measure of the influence of the past respect to the predator population xi. So, to smaller αi, longer is the interval in the past in which the values of S are taken into account (Cushing, 1977; MacDonald, 1978). This kernel has the property that
.
The behavior of the solutions of this system of ordinary differential equations will be analyzed looking to answer under what conditions will neither, one, or both species of predators survive and give some insight from the biological point of view. The adequate space for the initial data of the problem and some related notations is as follow:
Let denote the Banach space of the\ bounded and continuous function mapping from the interval to , the 3-dimensional vectors with positive coordinates. From the general theory of integral differential equations (Burton, 2005; Miller, 1971), for any initial data , there exists a unique solution for all t>0 and . Throughout, denote by the solution with , when no confusion arises. By a positive solution or of the previous integro-differential system, means that the solution has initial condition and each component of the solution is positive for all t>0. However, the model given by the previous integro-differential system can be associated to a system ordinary differential equations in the following way:
Introducing two new unknown functions, yi , defined by:
, (3)
i=1, 2 and using the form of the weak kernel (2) and the linear trick chain technique (MacDonald, 1978) gives the new system of ordinary differential equations:
(4)
Thus, the integro-differential system is “equivalent” to this system of 5-dimensional ordinary differential equations. The relationship between both systems are interpreted as follows: If is the solution of the integro-differential system corresponding to the continuous and bounded initial functions , then is solution of (4) with the initial conditions , with:
Conversely, if is any solution of (4), defined on the entire real line and bound ed on (−∞, 0], then yi , i=1, 2, is given by (2), so (S, x1 , x2 ) satisfies (3).
Most of the results of the paper are established considering the “equivalent system’’ (4).
3. Basic properties
In this section the basic properties of the model are established. In doing that, the following results are important for the proofs of some lemmas and theorems in the following. The first one is a lemma due to Barbalat and the proof may be seen in Gopalsamy (1992).
Lemma 1 (Barbălat lemma). Let a be a finite number and be a differentiable function. If exists (finite) and the derivative function g is uniformly continuous on (a, ∞), then .
The next definition and the theorem are due to Markus (1956) and will be needed for the proofs of some theorem in the next section.
Definition 1 Let and be a first order system of ordinary differential equations. The real-valued functions fi(x, t) and fi (x) are continuous in (x, t) for x ∈ G, where G is an open set in , and for t>t0, and they satisfy a local Lipschitz condition in x. A is said to be asymptotic to in G if for each compact set K ⊂ G and for each ε>0, there is a T = T(K, ε)>t0 such that:
for all , all and all
In relation with the previous definition, the Markus theorem follows.
Theorem 2 (Markus) Let in G and let P be an asymptotically stable critical point of A∞. Then there is neighborhood N of P and a time T such that the omega limit set for each solution x(t) of A, which intersects N at a time later than T is equal to P.
In the following results, the well definiteness of the system (4) is described. Note that the Barbălat Lemma implies that if , satisfies the system (4); then, according with the remark of Wolkowicz et al. (1997, p. 1288), each component of this solution is uniformly continuous.
The following preliminary two lemmas are basic for the well biological meaning of the integro-differential model. The first one indicates that the model possesses the property that positive initial data yield positive solutions.
Lemma 3 (Positivity) For any with , the solution remains positive for all t>0.
Proof. Clearly,
,
and so S(t)>0 for all t>0 since S(0)>0. To show that for all t>0 , suppose that it is not true. Let:
and ,
Then and . But from the integro-differential system, the following calculation gives:
This is a contradiction. Therefore, for any positive t. This completes the proof.
Note that the previous lemma implies that the set:
is invariant under the flow induced by the system (4). The following, second lemma, has to do with the property of pointwise dissipativity.
Lemma 4 (Pointwise Dissipativity) All positive solutions of model given by the integro-differential system are bounded for t>0. Moreover, system (4) is pointwise dissipative and the absorbing set B (into which every solution eventually enters and remains) is given by:
(5)
where
and
Proof. Of the equation for S in (4) it is easy to see that using a similar argument, as in the proof of Lemma 3.1 in Hsu et al. (1978b), the boundedness of S(t) can be obtained. Precisely, for sufficiently small ε>0 there exists T depending only on S(0), such that S(t))<K + ε , for t>T.
Now let:
then, for t>T,
where p = min {1, α1, α2}. And so,
W’ < K − pW.
This clearly implies the uniform boundedness of y1 and y2. Also note that taking into account the equations for xi in the equations (4), the boundedness of yi (t) implies the boundedness of xi (t). Taking into account the previous differential inequality and the equations for xi in the system (4), the number T1 = T1 (ε, P0 ) is obtained for P0 = (S0, x10, x20, y10, y20 ), in such a way that:
where , for all t>T1 . Thus, the pointwise dissipativity is obtained for the systems (4). This concludes the proof.
4. Properties of the equilibrium points
The following results shall give a complete description of the global asymptotic behavior of the system (4) under the generic condition μ1 ≠ μ2 , where the parameter μi is the ratio of the i−th predator’s Michaelis-Menten (half-saturation) constant to its intrinsic rate of increase, times is death rate, i.e.,
In this case, the equilibria of the system (4) are given by:
where:
The following result has to do with the inadequate predator, this result characterize the conditions under which the predators cannot survive on the prey, given the carrying capacity of the prey population, even in the absence of competition.
Lemma 5 A necessary condition for either species xi survive is 0<μi < K.
Proof. If S(t) ≥ K, then S(t)<0. Therefore, taking into account the absorbing set B given in (5), suppose that S(t)<K. Now let wi (t) = xi (t) + yi (t)/ αi , i = 1, 2. Then:
If mi ≤ Di , then the following representation is obtained:
And if μi ≥ K, then the following representation is obtained:
In any case, the appropriate representation implies that the positive function w(t) is decreasing and the limit of w(t) as t goes to infinity there exists. If , then by virtue of the uniform continuity of the derivatives of w(t) and the Barbălat lemma, . Therefore, if , then there exists a sequence , such that , and, therefore, . This contradicts the fact that S(t) < K < μi . Thus . That concludes the proof.
The previous lemma states that, if the maximum birth rate mi is less than or equal to the death rate Di , or if the parameter μi is greater or equal to the carrying capacity of the prey, then the ith predator will die out. This establishes that there is a minimum population size which can support a given predator: K must be larger than μi for the i−th predator survive, independent of competition. Thus, the Lemma 5 implies the following global result that describes the outcome in which both predator populations are eliminated because of an inadequate environment for either population to survive, rather than as a result of competition.
Theorem 6. If (a) m1 ≤ D1 or μ1 ≥ K, and (b) m2 ≤ D2 or μ2 ≥ K, then and .
Proof. Clearly the hypothesis implies that , and this implies that . With this, the remainder of the proof follows applying the Markus theorem in the same way as in the proof of Theorem 3.3(i) of Hsu et al. (1978b).
In the remainder of the paper is assumed that 0<μ1 < K.
Theorem 7. Let (a)
(8)
(9)
(10)
and (b) m2 ≤ D2 or μ2 ≥ K, then the equilibrium point E1 of the system (4) is globally asymptotically stable.
Proof. Clearly the hypothesis (b) implies that , and this implies that . In applying the Markus theorem let the system A be the system (4), and the system A∞ be given by:
Clearly A→A∞ in E. By other hand, the Jacobian matrix evaluated in the equilibrium point (μ1 , f1 (μ1), 0, D1 f1 (μ1), 0) of the system A∞ is given by:
and the characteristic polynomial of this matrix is given by newline:
Where:
The hypothesis (9) of the theorem implies that the two roots of the polynomial p1 have negative real part, and the hypothesis (8) implies that A<0 and B>0. Thus the Routh-Hurwitz criterion implies that all of the root of the polynomial p2 have negative real part. Because if were,
then:
Taking into account (8), this inequality implies that:
after some manipulations and taking into account (10) result that K>2μ1 + a1 , what is contradictory with (8). Thus, the equilibrium point (μ1 , f1 (μ1 ), 0, D1f1 (μ1 ), 0) of the system A∞ is asymptotically stable and by virtue of Markus lemma the conclusion of the theorem is obtained.
The following theorem guarantees the persistence of the species S, x1 , x2 under the necessary condition given in the previous lemma. Here the theory of uniform persistence used here is as appear in Thieme (1993).
Theorem 8. If 0<μi < K, i ∈ {1, 2}, then the all the populations S, x1 , x2 are persistent.
5. The one predator-one prey system
In order to have a comprehensive insight of the model, in this section the analysis of the one predator-one prey system is presented. In this case, the system (4) reduces to the three-dimensional system of ordinary differential given by:
Denote by 𝒜 the open region and define by μ, the lumping parameter given by μ = aD/m−D. It is easy to see that the only equilibrium points of system (12) in the boundary of 𝒜 are P0 = (0, 0, 0), which is unstable, and PK = (K, 0, 0), which is globally asymptotically for μ>K. There are not equilibrium points with positive coordinates for the previous mentioned values of the parameters. When μ = K, the equilibrium PK loses his stability (see Table 1) and a saddle-node bifurcation arises and for μ<K appears the only equilibrium, P* in the positive orthant, which is given by:
, where
Letting u = (S, x, y)T, v = (S, x, z)T, and H=diag [1, 1, −1]. The transformation v = Hu takes the system (12) into a competitive and irreducible system in the sense as appears in Smith (2008). Note that system (12) may be written as:
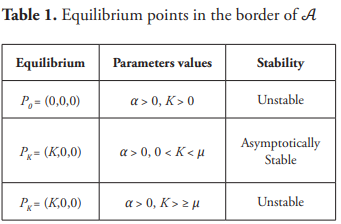
where h(S) = mS/a+S. With these notations, the Jacobian matrix of system (13) evaluated at the equilibrium point P* takes the form:
and the characteristic polynomial of the matrix A is given by:
where,
To see the sign of the real part of the roots of the polynomial p(λ), the Routh-Hurwitz criterion will be used. Clearly, if K>a + 2μ, then the equilibrium point P* is unstable since the coefficient b1 is negative. Therefore, the stability of the equilibrium point P* requires filling the condition:
(14)
The Routh-Hurwitz criterion implies that negative the real part of the roots of p(λ) occurs if and only if the following inequality hold:
(15)
where G = (D/mK) (a + 2μ − K) and L = aD2 /mKμ (K − μ). From this inequality the following polynomial is obtained:
Where the parameter K is emphasized as an argument of the polynomial q in order to explain better his sign as α varies respect to K. The inequality (15) holds, if and only if, q(α, K)>0. The sign of the polynomial q(α, K) depends on the coefficient of the term α + D. By other hand, the function q has a minimum respect to α at α* + D = − 1/2 r(K)/G, given by:
(16)
When this minimum is positive, then clearly q(α, K)>0 and the equilibrium point is stable for all K>μ. Now, putting:
(17)
and note that r(μ) is positive, and r(a + 2μ) is negative, which implies that there exists a unique value of K between μ and a + 2μ, say K* , such that r(K*) = 0. Thus, for K in the interval (μ, K* ], r(K) ≥ 0, and therefore the coefficient (17) of the term α+D of the polynomial q(α, K) is positive, which implies that q(α, K)>0, for all positive α, and in this case the equilibrium point P* is (locally) asymptotically stable. When K ∈ (K* , a + 2μ), the sign of r(K) reversed and in this case the coefficient of the term α+D of the polynomial q(α, K) (17) is negative. In this case, q(α, K) has two zeros α1 + D, α2 + D, such that q(α, K) ≤ 0 for α ∈ (α1 + D, α2 + D) and the equilibrium point P* is unstable. Note that all of this situation occurs when the minimum of q(α, K) is negative, and clearly:
(18)
All of the previous calculations are summarized in the following theorem. The proof of the Hopf bifurcation isnvolve several calculations and the procedure is similar to the used in in Cavani & Farkas (1994).
Theorem 9. A necessary condition for the existence of the equilibrium of positive coordinates P* of the system (12) is μ<K. Moreover: (i) If μ<K<a + 2μ, then there exists a unique number K* between μ and a + 2μ such that if K belongs to the interval (μ, K* ), the equilibrium P* is (locally) asymptotically stable for all positive α. When K = K * , the equilibrium P* is a center (stable). For K>K* the equilibrium point P* , lost its stability and for K in the interval (K* , a + 2μ) there are two values of α, α1 and α2 such that for α ∈ (α1 + D, α2 + D), q(α, K) in (16) is negative and therefore the equilibrium P* is unstable. If K = K* and α = α1 + D, a transcritical Hopf bifurcation takes place. Finally, for K ≥ a + 2μ the coefficient b1 of the characteristic polynomial p(λ) of the matrix A is negative and therefore the equilibrium point P* is unstable.
Note from (17) that for K<K* very near to the value K* the term r(K) is positive and the equilibrium point P* goes from to be stable to unstable as K passes through K* and α through α1 + D . This can be interpreted as the system (13) exhibits the paradox of the enrichment; however, the delay may act as a mechanism to regain the stability. In fact, when 1/α increases and passes through 1/α1 +D, the equilibrium point P* is again stable. The Table 2 that follows may be useful in order to get in mind the stability of the equilibrium P* .
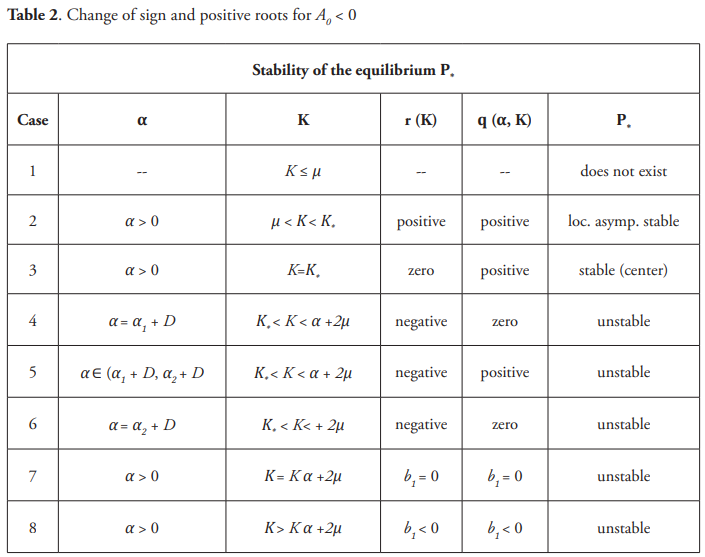
Example. For the following values of the parameters: m = 2, a = 0.5, D = 1, give μ = 0.5, a + 2μ = 13.5, K* = 1.18. and picking K = 1.17, then for α = 0.9 the equilibrium point P* is (locally) asymptotically stable. For K = 2 and α = 0.9 there is a stable periodic orbit.
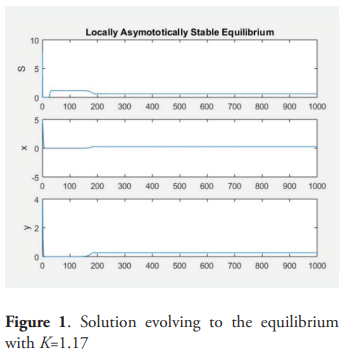
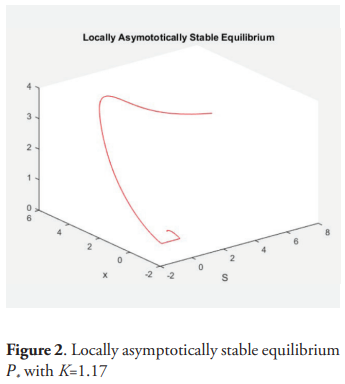
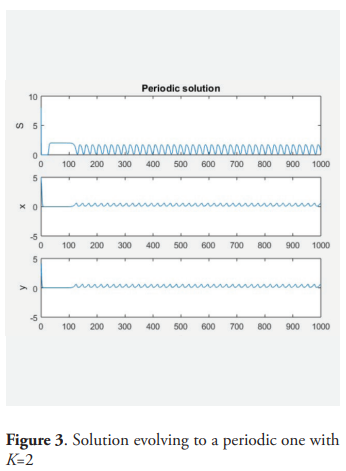
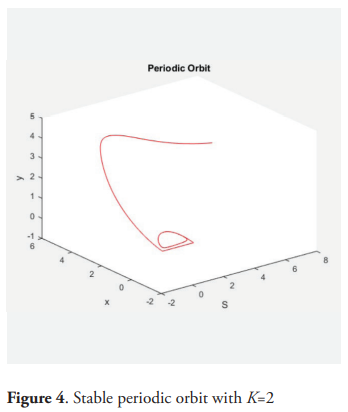
6. Conclusions
This paper is an analysis of the behavior of a model of two predators competing exploitatively for a shared prey species. The prey grows logistically in absence of predation and the predators consume prey according to a saturating functional response, but the growth of the predator is influenced by the amount of prey in the past. It is supposed that the predator grows up depending on the weight average time of the function of Michaelis-Menten of S over the past per predator. The system is equivalent to a dissipative system of five ordinary differential equation, by using the so-called trick of the linear chain. The analysis of this equivalent system establishes that the species survive if and only if K>μi, where K is the carrying capacity of the prey specie and μi is the ratio of the i−th predator according with the Michaelis-Menten response. The case when there exists only one predator is analyzed with detail. In this case the equilibrium point of positive coordinates is unstable for all K>a + 2μ. Theorem 9 characterizes the general situation and the occurrence of the Hopf bifurcation. The table 2 describes all of the situations analyzed and the figures 1, 2, 3 and 4 give examples of the dynamic.
Referencias
Burton, T. (2005). Volterra Integral and Differential Equations, Volume 202, Second Edition (Mathematics in Science and Engineering). Amsterdam: Elsevier.
Cavani, M. & Farkas, M. (1994). Bifurcations in predator-prey model with memory and diffusion. I: Andronov-Hopf bifurcation. Acta Mathematica Hungarica, 63, 213-229.
Cavani, M., Lizana, M., & Smith, H. L. (2000). Stable periodic orbits for a predator-prey model with delay. Journal of Mathematical Analysis and Applications, 249, 324-339.
Cushing, J. M. (1977). Integrodifferential Equations and Delay Models in Population Dynamics 20, Heidelberg: Springer-Verlag.
Gopalsamy, K. (1992). Stability and Oscillations in Delay Differential Equations of Populations. Dynamics The Netherlands: Kluwer, Dordrecht.
Hsu, S. B. (1978). Limiting behavior for competing species. SIAM Journal on Applied Mathematics. 34, 760-763.
Hsu, S. B., Hubbell, S. P., & Waltman, P. (1977). A mathematical theory for single-nutrient competition in continuous culture of micro-organisms. SIAM Journal on Applied Mathematics. 32, 366-383.
Hsu, S. B., Hubbell, S. P., & Waltman, P. (1978a). A contribution to the theory of Competing Predators. Ecological Monographs, 48, 337-349.
Hsu, S. B., Hubbell, S. P., & Waltman, P. (1978b). Competing Predators. SIAM Journal on Applied Mathematics. 35, 617-625.
MacDonald, N. (1978). Time Lags in Biological Models, Lecture Notes in Bio-mathematics, 27, Heidelberg: Springer-Verlag.
Markus, L. (1956). Asymptotically autonomous differential systems, Contribution to the Theory of Nonlinear Oscillations 3. New Jersey: Princeton University Press.
Miller, R. K. (1971). Nonlinear Volterra Integral Equations. W. A. Benjamin (ed.). California: Menlon Park.
Smith, H. L. (2008). Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems: An Introduction to the Theory of Competitive and Cooperative Systems. New York: American Math. Soc.
Thieme, H. R. (1993). Persistence under relaxed point-dissipativity (with application to an epidemic model). SIAM Journal on Applied Mathematics, 24, 407-435.
Wolkowicz, G., H. Xia, H., & Ruan, S. (1997). Competition in the Chemostat: A distributed delay model and its global asymptotic behavior. SIAM Journal on Applied Mathematics, 57, 1281-1310.
